Показатель VO2max считается одним из самых общедоступных (он есть в продвинутых спортивных часах), но и одним из самых непонятных для любителя бега. Зачем он нужен, откуда он берется, стоит ли использовать его как ориентир для отслеживания прогресса? Попробуем разобраться.
Бегуны всех уровней, от увлеченных любителей до профессионалов, ищут пути для повышения эффективности тренировок для улучшения результатов и установления новых рекордов. Бег на длинные дистанции требует от участников большого объема тренировок на выносливость для преодоления постоянного физиологического стресса. Различные способы манипуляции физиологическими параметрами для улучшения выносливости и эффективности бегунов ведутся вот уже более 30 лет, хотя остается достаточное количество вопросов (1). Большинство методик, известных сегодня, появились в результате многочисленных проб и ошибок, а чёткое научное обоснование получили лишь некоторые из них (2, 3, 4).
Длительное время показатель максимального потребления кислорода (VO2max) используется в качестве некоей «магической пули», позволяя выстраивать тренировки на основании его значения и проводить анализ производительности и прогресса атлета. Но так ли он хорош, всем ли подходит и можно ли на него полагаться?
Считается, что для каждого увлеченного бегом человека, показатель VO2max (или VDOT у Дэниелса) фактически определяет его талант или потенциал. Величина VO2max определяет максимальное потребление кислорода, и это один из наиболее часто используемых показателей для отслеживания прогресса в тренировках.
Конечно, все мы слышали про невероятные цифры VO2max у многих профессиональных спортсменов: Lance Armstrong (84 мл/кг/мин), Steve Prefontaine (84,4 мл/кг/мин), Bjørn Dæhlie (96 мл/кг/мин) и многих других. Но нужно ли уделять такое пристальное внимание этим цифрам? Если говорить вкратце, то нет.
В противоположность бытующему мнению, VO2max – это просто измерение, оно не характеризует тренированность или потенциал атлета. Фактически, среди нескольких тренированных бегунов невозможно определить быстрейшего, основываясь только на показателе VO2max. Измерение VO2max не очень точно отражает важнейшие процессы транспорта и утилизации кислорода в мышцах. Попробуем для начала внимательно рассмотреть этот показатель, его составляющие, а также влияние, которые различные этапы транспорта кислорода оказывают на VO2max.
КОНЦЕПЦИЯ VO2max.
Термин «максимальное потребление кислорода» впервые был описан и использован Hill (5) и Herbst (6) в 1920-х годах (7). Основные положения теории VO2max гласили:
1. Существует верхняя граница потребления кислорода;
2. Существует естественная разница в значениях VO2max;
3. Высокий VO2max необходим для успешного участия в забегах на средние и длинные дистанции;
4. VO2max ограничен способностью сердечно-сосудистой системы переносить кислород к мышцам.
Показатель VO2max характеризует максимальное количество используемого кислорода, и рассчитывается путем вычитания количества выдохнутого кислорода из количества поглощенного кислорода (8). Поскольку VO2max используется для количественного описания ёмкости аэробной системы, показатель находится под влиянием большого количества факторов на длинном пути кислорода от окружающей среды до митохондрий в мышцах.
Формула для расчета VO2max: VO2max= Q х (CaO2-CvO2),
Где Q – сердечный выброс, CaO2 – содержание кислорода в артериальной крови, CvO2 - содержание кислорода в венозной крови.
Это уравнение принимает в расчет объем крови, перекачиваемый нашим сердцем (сердечный выброс = ударный объем х частота сердечных сокращений), а также разницу между уровнем кислорода в крови, притекающей в мышцы (CaO2 – содержание кислорода в артериальной крови) и уровнем кислорода в крови, оттекающей от мышц к сердцу и лёгким(CvO2 - содержание кислорода в венозной крови). По сути, разница (CaO2-CvO2) представляет собой количество кислорода, поглощенного мышцами. Хотя для практических целей измерение VO2max имеет небольшое значение, развитие способности более эффективно потреблять и утилизировать кислород влияет на производительность бегуна.
Поглощение и утилизация кислорода, в свою очередь, зависят от целого ряда факторов, которые встречаются на длинном пути кислорода. Движение кислорода от атмосферного воздуха до митохондрий называется кислородным каскадом. Вот его основные этапы:
1. Потребление кислорода:
- Поступление воздуха в лёгкие;
- Движение по трахеобронхиальному дереву до альвеол и капилляров, где кислород поступает в кровь.
2. Транспорт кислорода:
- Сердечный выброс – кровь поступает к органам и тканям;
- Концентрация гемоглобина;
- Объем крови;
- Капилляры, из которых кислород поступает в мышцы.
3. Утилизация кислорода:
- Транспорт в митохондрии;
- Использование в аэробном окислении и цепи переноса электронов.
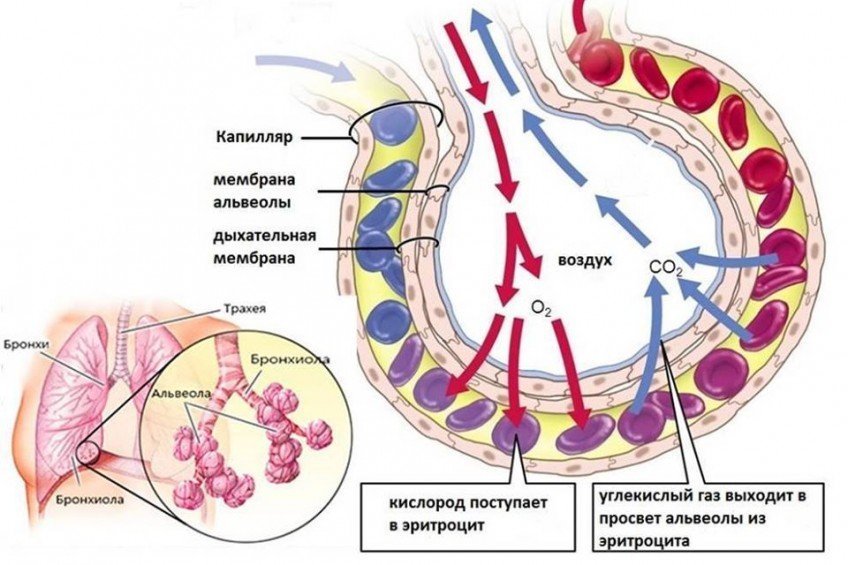
ПОТРЕБЛЕНИЕ КИСЛОРОДА.
Первый этап путешествия кислорода состоит в его поступлении в лёгкие и в кровоток. За эту часть, в основном, отвечает наша дыхательная система. Воздух попадает из ротовой и носовой полости в лёгкие благодаря разнице давлений между лёгкими и внешней средой (во внешней среде давление кислорода больше, чем в лёгких, и кислород «засасывается» внутрь наших лёгких). В лёгких воздух движется по бронхам к более мелким структурам, называемым бронхиолы. На конце бронхиол есть специальные образования - дыхательные мешочки, или альвеолы.
Альвеолы – это место переноса (диффузии) кислорода из лёгких в кровь, а точнее в капилляры, оплетающие альвеолы (Представьте себе шарик, опутанный паутиной – это и будут альвеолы с капиллярами). Капилляры - самые мелкие кровеносные сосуды в организме, их диаметр равен всего 3-4 микрометра, это меньше диаметра эритроцита. Получая кислород из альвеол, капилляры затем несут его в более крупные сосуды, которые в конечном итоге впадают в сердце. Из сердца по артериям кислород разносится во все ткани и органы нашего тела, в том числе и мышцы.
Количество поступающего в капилляры кислорода зависит как от наличия разницы давлений между альвеолами и капиллярами (содержание кислорода в альвеолах больше, чем в капиллярах), так и от общего количества капилляров. Количество капилляров играет определенную роль, особенно у хорошо тренированных атлетов, поскольку позволяет большему объему крови протекать через альвеолы, способствуя поступлению большего количества кислорода в кровь.
Использование или потребность в кислороде зависит от скорости бега. При повышении скорости, большее количество клеток в мышцах ног становится активно, мышцам необходимо больше энергии для поддержания проталкивающего движения, а значит, мышцы потребляют кислород с более высокой скоростью. Фактически, потребление кислорода линейно связано со скоростью бега (выше скорость – больше кислорода потребляется.
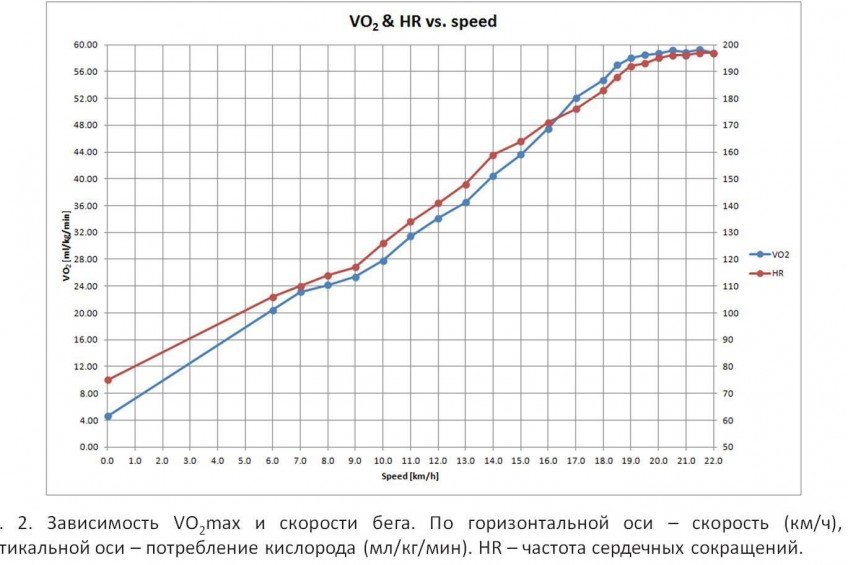
Средний бегун, развивающий скорость 15 км/ч, скорее всего, будет потреблять кислород со скоростью 50 мл на килограмм веса в минуту (мл/кг/мин). При 17,5 км/ч, скорость потребления вырастет почти до 60 мл/кг/мин. Если бегун способен развить скорость 20 км/ч, потребление кислорода будет еще выше – около 70 мл/кг/мин.
Тем не менее, показатель VO2max не может расти бесконечно. В своем исследовании Hill описывает ряд изменений VO2 у атлета бегущего по травяному треку с разной скоростью (9). После 2.5 минут бега на скорости 282 м/мин, его VO2 достиг значения 4.080 л/мин (или 3.730 л/мин выше измеренного значения в покое). Поскольку VO2 при скоростях 259, 267, 271 и 282 м/мин не возрастал выше значения полученного при скорости бега 243 м/мин, это подтвердило предположение, что при высоких скоростях VO2 достигает максимума (плато), превысить который невозможно, как бы ни увеличилась скорость бега.
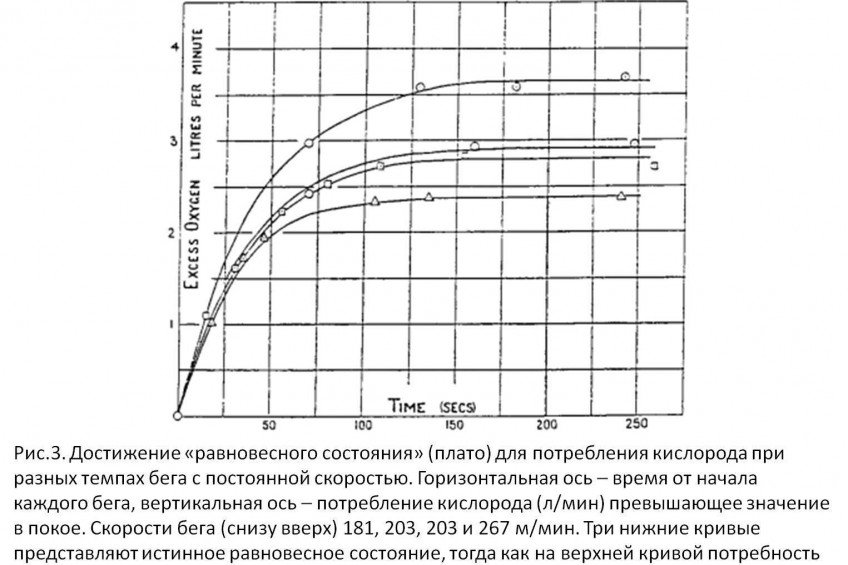
Сегодня общепринят факт существования физиологической верхней границы возможностей организма потреблять кислород. Это наилучшим образом было проиллюстрировано на классическом графике Åstrand и Saltin (10).
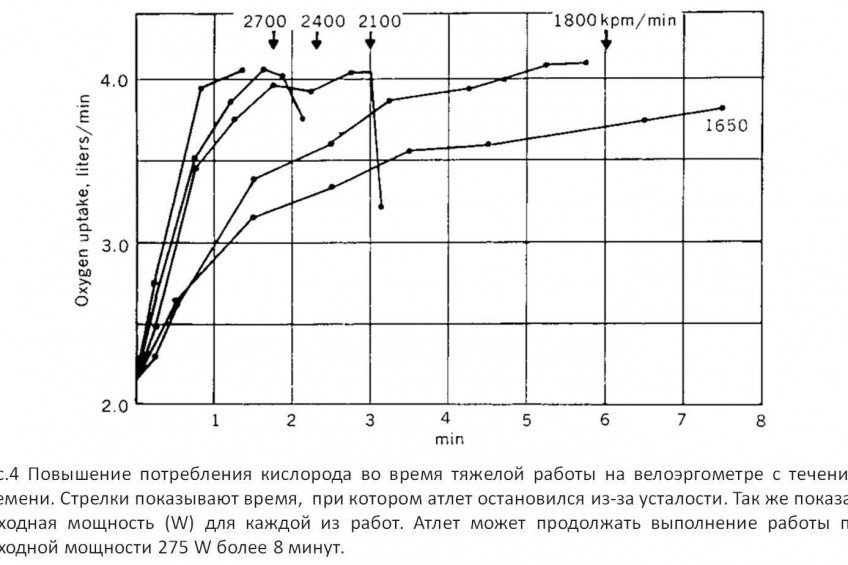
Говоря про интенсивность работы, необходимо уточнить один факт. Даже при высокой интенсивности, насыщение крови кислородом не падает ниже 95% (это на 1-3% ниже показателя здорового человека в состоянии покоя). Этот факт используется как показатель того, потребление и транспорт кислорода из легких в кровь не является ограничивающими факторами производительности, поскольку насыщение крови остается высоким.
Однако у некоторых тренированных атлетов описан феномен, известный как «артериальная гипоксемия (гипоксемия – низкий уровень кислорода в крови, кислородное голодание), вызванная физической нагрузкой» (11). Это состояние характеризуется падением насыщения кислорода на 15% при выполнении упражнений, относительно уровня покоя. Падение кислорода на 1% при насыщении кислорода ниже 95% приводит к снижению VO2max на 1-2% (12). Причина развития этого феномена следующая. Высокий сердечный выброс тренированного атлета приводит к ускорению кровотока через лёгкие, и кислород попросту не успевает насытить протекающие через лёгкие кровь. Для аналогии, представьте поезд, проходящий через небольшой городок в Индии, где люди часто запрыгивают в поезда на ходу. При скорости поезда 20 км/ч в поезд смогут запрыгнуть, скажем, 30 человек, тогда как при скорости поезда 60 км/ч, в него запрыгнут 2-3 человека в лучшем случае. Поезд – это сердечный выброс, скорость поезда – это кровоток через лёгкие, пассажиры – это кислород, старающийся попасть из лёгких в кровь. Таким образом, у некоторых тренированных атлетов, потребление и диффузия кислорода из альвеол в кровь все-таки может влиять на величину VO2max.
Помимо диффузии, сердечного выброса, количества капилляров, на VO2max и насыщение крови кислородом может влиять сам процесс дыхания, точнее мышцы, участвующие в процессе дыхания. Так называемая «кислородная цена» дыхания оказывает значимое влияние на VO2max. У «обычных» людей при умеренно интенсивной физической активности на дыхание тратится примерно 3-5% от поглощенного кислорода, а при высокой интенсивности эти затраты вырастают до 10% от величины VO2max (13).
Другими словами, на процесс дыхания (работу дыхательных мышц) затрачивается какая-то часть от поглощенного кислорода. У тренированных атлетов в ходе интенсивных нагрузок на дыхание тратится 15-16% от VO2max (14). Более высокая цена дыхания у хорошо тренированных атлетов, подтверждает предположение о том, что потребность в кислороде и факторы, ограничивающие производительность у тренированных и нетренированных людей разные.
Другая возможная причина того, что процесс дыхания может ограничивать производительность атлета, это существующая «конкуренция» за кровоток между дыхательными мышцами (в основном диафрагмой) и скелетными мышцами (например, мышцы ног). Грубо говоря, диафрагма может «оттягивать» на себя часть крови, которая не попадает из-за этого в мышцы ног. Из-за такого соперничества, усталость диафрагмы может произойти при уровне интенсивности выше 80% от VO2max (15).
Другими словами, при условно-средней интенсивности бега, диафрагма может «устать» и работать менее эффективно, что приводит к обеднению организма кислородом (поскольку диафрагма отвечает за вдох, при усталости диафрагмы его эффективность снижается, и лёгкие начинаю работать хуже).
В проведенном обзоре Sheel и соавторы показали, что после включения в тренировочный цикл специальных дыхательных упражнений, атлеты показали улучшение производительности (16). Эту гипотезу подтвердило исследование проведенное на велосипедистах, когда во время 20 и 40 километровых отрезков у спортсменов развивалась глобальная усталость мышц вдоха (17). После тренировки дыхательных мышц у атлетов было обнаружено улучшение производительности на 20 и 40 километровых отрезков на 3,8% и 4,6%, соответственно, а также уменьшение усталости дыхательных мышц после отрезков.
Таким образом, дыхательные мышцы влияют на VO2max, причем это степень этого влияния зависит от уровня тренированности. Для атлетов более высокого уровня важными ограничивающими факторами будут утомление дыхательных мышц и гипоксемия (недостаток кислорода), вызванная физической активностью. В связи с этим, хорошо тренированные спортсмены должны использовать дыхательную тренировку, тогда как бегуны начального уровня, скорее всего, не получат от нее такого же эффекта.
Самым простым способом тренировки дыхательных мышц, применяющимся и в клиниках, является выдох через неплотно сжатые губы. Необходимо почувствовать, что выдыхаешь всей диафрагмой, начать с медленных и глубоких вдоха и выдоха, постепенно наращивая скорость выдоха.
ТРАНСПОРТ КИСЛОРОДА.
Со времен первых экспериментов A.V. Hill по измерению VO2max, транспорт кислорода всегда считался главным ограничивающим фактором для показателя VO2max (18).Было подсчитано, что транспорт кислорода (это весь путь от поступления кислорода в кровь до его поглощения мышцами) влияет на VO2max примерно на 70-75% (19). Одним из важных компонентов транспорта кислорода является его доставка к органам и тканям, которая также подвержена влиянию большого количества факторов.
АДАПТАЦИЯ СЕРДЕЧНО-СОСУДИСТОЙ СИСТЕМЫ.
Сердечный выброс (СВ) – это количество крови, выбрасываемой сердцем в минуту, он также считается важным фактором, ограничивающим VO2max. Сердечный выброс зависим от двух факторов – частоты сердечных сокращений (ЧСС) и ударного объема (УО). Следовательно, для увеличения максимального СВ, один из этих факторов должен быть изменен. Максимальная ЧСС не меняется под влиянием тренировок на выносливость, тогда как УО у спортсменов повышается как в состоянии покоя, так и при выполнении работы любой интенсивности. Повышение УО происходит за счет увеличения размеров и сократимости сердца (20). Эти изменения в сердце вызывают улучшение способности быстро заполнять камеры сердца.
Согласно закону Франка-Старлинга, при увеличении растяжения камеры сердца перед сокращением, само сокращение будет более сильным. Для аналогии можно представить себе полоску резины, которую растягивают. Сильнее растяжение – быстрее сокращение. Это означает, что заполнения камер сердца у атлетов вызовет более быстрое сокращение сердца, а значит, приведет к увеличению ударного объема. В дополнение к этому, у бегунов на длинные дистанции появляется способность быстро заполнять камеры сердца при высокой интенсивности нагрузки. Это достаточно важное физиологическое изменение, поскольку в норме при увеличении частоты сердечных сокращений остается меньше времени на заполнение камер сердца.
ГЕМОГЛОБИН.
Другим важным фактором в транспорте кислорода является способность крови переносить кислород. Эта способность зависит от массы красных кровяных телец, эритроцитов, а также концентрации гемоглобина, который служит основным переносчиком кислорода в организме.
Повышение гемоглобина должно улучшить производительность благодаря повышению транспорта кислорода к мышцам. Исследования четко показывают эту взаимосвязь, изучая, как снижение уровня гемоглобина повлияет на производительность (21). Например, снижение уровня гемоглобина при анемии приводит к снижению VO2max (22). Так, в одном из исследований после снижения уровня гемоглобина наблюдалось снижение VO2max, гематокрита и выносливости. Однако после двух недель было отмечено восстановление начального значения VO2max, а гемоглобин и выносливость оставались сниженными (23). Факт сохранения нормальных значений VO2max может при низком уровне гемоглобина поднимает ряд вопросов и демонстрирует обширные адаптационные возможности организма, напоминая о том, что существует огромное количество способов оптимизировать доставку кислорода для повышения VO2max. Кроме того, возвращение VO2max, но не выносливости, к нормальным показателям, может говорить о том, что VO2max и выносливость не являются синонимами.
На другом конце спектра исследования, где искусственно повышался уровень гемоглобина. Эти работы показали повышение как VO2max, так и производительности (24). Одиннадцать элитных бегунов, включенных в одно из исследований, продемонстрировали значительное удлинение времени до момента наступления истощения и VO2max после переливания крови и повышения уровня гемоглобина со 157 г/л до 167 г/л (25). В исследовании с кровяным допингом, который приводит к искусственному повышению гемоглобина, отмечалось улучшение VO2max на 4% - 9% (Gledhill 1982). Соединенные воедино, все вышеперечисленные факты свидетельствуют о том, что уровень гемоглобина оказывает значимое влияние на VO2max.
ОБЪЕМ КРОВИ.
С повышением гемоглобина кровь становится более вязкой, поскольку большая часть крови содержит эритроциты, а не плазму. При повышении количества эритроцитов увеличивается вязкость и растет такой показатель, как гематокрит. Для аналоги, представьте себе, как текут по трубам одно и того же диаметра вода (это аналог крови с нормальным гемоглобином и гематокритом) и кисель (гемоглобин и гематокрит повышен).
Гематокрит определяет отношение между эритроцитами и плазмой. При высокой вязкости крови кровоток замедляется, затрудняя, а иногда и полностью прекращая доставку кислорода и нутриентов к органам и тканям. Причина – кровь с высокой вязкостью очень «лениво» течет, а в самые маленькие сосуды, капилляры, может и не попасть, попросту закупоривая их. Следовательно, чересчур высокий гематокрит может потенциально снизить производительность через нарушение доставки кислорода и нутриентов к тканям.
При тренировках на выносливость нормальной ситуацией является повышение как объема крови, так и гематокрита с гемоглобином, причем увеличение объема крови может доходить до 10% (26). В медицине достаточно много раз менялась концепция так называемого оптимального гематокрита, и до сих пор не утихают споры, какой же уровень этого показателя считать оптимальным. Очевидно, что однозначного ответа на этот вопрос не существует, и для каждого атлета уровень гематокрита, при котором есть максимальная выносливость и работоспособность можно считать оптимальным.
Однако необходимо помнить, что высокий гематокрит - это не всегда хорошо.
Атлеты, использующие запрещенные препараты, например эритропоэтин (ЭПО) для искусственного повышения уровня эритроцитов будут отличаться очень хорошей выносливостью и работоспособностью. Обратной стороной медали при этом может являться опасно высокий уровень гематокрита, а также повышение вязкости крови (27). С другой стороны, есть атлеты с хорошей выносливостью, которые бегают с низким уровнем гематокрита и гемоглобина, что в обычной жизни может быть признаком анемии. Вполне возможно, что подобные изменения являются ответом на высотную адаптацию спортсменов.
Адаптация к высокогорью может быть трех разных видов (28):
Эфиопия – поддержание баланса между насыщением крови и гемоглобином;
Анды – повышение уровня эритроцитов со снижением насыщения крови кислородом;
Тибет – нормальная концентрация гемоглобина со снижением насыщения крови кислородом.
Несколько вариантов адаптации говорят о том, что существует несколько способов оптимизировать показатели крови. Ответа и на вопрос, у кого же из вариантов (низкий или высокий гематокрит) в спорте лучше доставка кислорода, до сих пор нет. Скорее всего, как бы ни банально это прозвучало, ситуация с каждым атлетов индивидуальная.
Другим важнейшим параметром, играющим роль во время бега, является так называемое шунтирование крови. Этот механизм полезен, когда мышцам необходимо больше крови и кислорода с нутриентами. Если в покое скелетная мускулатура получает только 15-20% от общего объема крови, то при интенсивной физической нагрузке примерно 80-85% от общего объема крови идут к мышцам. Процесс регулируется расслаблением и сокращением артерий. Кроме того, при тренировках на выносливость повышается плотность капилляров, по которым все необходимые вещества поступают в кровь. Доказано также, что плотность капилляров напрямую связана с VO2max (29).
УТИЛИЗАЦИЯ КИСЛОРОДА.
Как только кислород поступил к мышцам, он должен быть утилизирован. За утилизацию кислорода отвечают «энергетические станции» наших клеток – митохондрии, в которых кислород используется для производства энергии. О том, как много кислорода поглотили мышцы, можно судить по «артериовенозной разнице», то есть разнице между содержанием кислорода в притекающей (артериальной) к мышце крови и содержанием кислорода в оттекающей (венозной) от мышцы крови. Другими словами, если притекает 100 единиц кислорода, а оттекает 40, тогда артериовенозная разница составит 60 единиц – именно столько усвоилось мышцами.
Артериовенозная разница не является фактором, ограничивающим величину VO2max по ряду причин. Во-первых, эта разница достаточно схожа как у элитных бегунов, так и у непрофессионалов (30). Во-вторых, если посмотреть на артериовенозную разницу, то видно, что кислорода в вене остается очень немного. Содержание кислорода в крови, притекающей к мышцам примерно равняется 200 мл кислорода на 1 литр крови, а в оттекающей венозной крови кислорода содержится всего около 20-30 мл на литр крови (29). Интересно, что показатель артериовенозной разницы может улучшаться в ходе тренировок, что означает большее поглощение кислорода мышцами. В нескольких исследованиях было показано увеличение показателя артериовенозной разницы примерно на 11% под влиянием систематических тренировок на выносливость (31). Учитывая все эти факты, можно сказать, что хотя артериовенозная разница не является ограничивающим VO2max фактором, но во время тренировок на выносливость происходят важные и полезные изменения данного показателя, свидетельствующие о большем поглощении кислорода мышцами.
Кислород заканчивает свой длинный путь в митохондриях клетки. Митохондрии скелетной мускулатуры – это место выработки аэробной энергии. В самих митохондриях кислород участвует в цепи переноса электронов, или дыхательной цепи. Таким образом, количество митохондрий играет важную роль в генерации энергии. В теории, чем больше митохондрий, тем больше кислорода может утилизироваться в мышцах. Исследования показали, что количество митохондриальных ферментов увеличивается при тренировках, однако рост VO2max при этом небольшой. Роль митохондриальных ферментов заключается в усилении реакции в митохондриях, для значительного увеличения продукции энергии. В одном исследовании, изучавшем изменения во время и после прекращения тренировок, мощность митохондрий увеличивалась на 30% в ходе тренировок, тогда как VO2max повышался всего на 19%. Однако, после прекращения тренировок показатель VO2max сохранялся дольше, чем мощность митохондрий (32).
ВЫВОДЫ:
1. Показатель VO2max характеризует максимальное количество используемого кислорода;
2. VO2max используется для количественного описания ёмкости аэробной системы;
3. Для практических целей измерение VO2max имеет небольшое значение, однако развитие способности более эффективно потреблять и утилизировать кислород влияет на производительность бегуна;
4. При повышении скорости бега мышцы потребляют кислород с более высокой скоростью;
5. Для показателя VO2max есть конечная точка роста, после чего он выходит на плато, или равновесное состояние;
6. Сам процесс дыхания значимо влияет на VO2max;
7. Дыхательные мышцы влияют на VO2max, причем это степень этого влияния зависит от уровня тренированности;
8. Максимальная частота сердечных сокращений не меняется под влиянием тренировок на выносливость, тогда как ударный объем у спортсменов повышается как в состоянии покоя, так и при выполнении работы любой интенсивности;
9. Уровень гемоглобина оказывает значимое влияние на VO2max;
10. Чересчур высокий гематокрит может потенциально снизить производительность через нарушение доставки кислорода и нутриентов к тканям.
Автор материала Доктор Евгений Суборов.
СПИСОК ЛИТЕРАТУРЫ:
1. Pollock ML. The quantification of endurance training programs. Exerc Sport Sci Rev. 1973; 1: 155-88
2. Hawley JA. State of the art training guidelines for endurance performance. S Afr J Sports Med 1995; 2: 7-12
3. Hawley JA, Myburgh KH, Noakes TD, et al. Training tech niques to improve fatigue resistance and endurance perform ance. J Sports Sci 1997; 15: 325-33
4. Tabata I, Irisawa K, Kouzaki M, et al. Metabolic profile of high intensity intermittent exercises. Med Sci Sports Exerc 1997; 29: 390-5
5. A.V. Hill and H. Lupton. Muscular exercise, lactic acid, and the supply and utilization of oxygen. Q. J. Med. 16:135–171, 1923
6. R. Herbst. Der Gasstoffwechsel als Mass der korperlichen Leistungsfahigkeit. I. Mitteilung: die Bestimmung des Sauerstoffaufnahmevermogens bein Gesunden. Deut. Arch. Klin. Med. 162: 33–50, 1928
7. B. Saltin and S. Strange. Maximal oxygen uptake: “old” and “new” arguments for a cardiovascular limitation. Med. Sci. Sports Exerc. 24:30–37, 1992
8. D.R. Bassett Jr, E.T. Howley. Limiting factors for maximum oxygen uptake and determinants of en-durance performance. Med Sci Sports Exerc. 2000 Jan; 32(1):70-84.
9. A.V. Hill, C.N.H. Long, and H. Lupton. Muscular exercise, lactic acid and the supply and utilisation of oxygen: Parts VII-VIII. Proc. Roy. Soc. B 97:155–176, 1924.
10. P.O. Åstrand, and B. Saltin. Oxygen uptake during the first minutes of heavy muscular exercise. J. Appl. Physiol. 16:971–976, 1961.
11. S.K. Powers, J. Lawler, J.A. Dempsey, S. Dodd, G. Landry. Effects of incomplete pulmonary gas ex-change on VO2 max. J Appl Physiol. 1989 Jun; 66(6):2491-5.
12. J.A. Dempsey, P.D. Wagner. Exercise-induced arterial hypoxemia. J Appl Physiol. 1999 Dec; 87(6): 1997-2006
13. E.A. Aaron, K.C. Seow, B.D. Johnson, J.A. Dempsey. Oxygen cost of exercise hyperpnea: implications for performance. J Appl Physiol 1992; 72: 1818–1825.
14. C.S. Harms, T.J. Wetter, S.R. McClaran, D.F. Pegelow, G.A. Nickele, W.B. Nelson, P. Hanson, J.A. Dempsey. Effects of respiratory muscle work on cardiac output and its distribution during maximal exercise. J Appl Physiol. 1998; 85: 609–618.
15. B.D. Johnson, M.A. Babcock, O.E. Suman, J.A. Dempsey. Exerciseinduced diaphragmatic fatigue in healthy humans. J.Physiol 1993; 460; 385-405.
16. A.W. Sheel. Respiratory muscle training in healthy individuals: physiological rationale and implica-tion for exercise performance. Sports Med 2002; 32(9): 567-81
17. L. M. Romer, A. K. McConnell, D. A. Jones. Effects of inspiratory muscle training on time-trial per-formance in trained cyclists. Journal of Sports Sciences, 2002; 20: 547-562
18. D.R. Bassett Jr, E.T. Howley. Limiting factors for maximum oxygen uptake and determinants of en-durance performance. Med Sci Sports Exerc. 2000 Jan; 32(1):70-84.
19. P. E. di Prampero. Factors limiting maximal performance in humans. Eur J Appl Physiol. 2003; Oct; 90(3-4): 420-9.
20. G. C. Henderson, M. A. Horning, S. L. Lehman, E. E. Wolfel, B. C. Bergman, G. A. Brooks. Pyruvate shuttling during rest and exercise before and after endurance training in men. Journal of Applied Physiology Jul 2004; 97(1): 317-325
21. J.A. Calbet, C. Lundby, M. Koskolou, R. Boushel. Importance of hemoglobin concentration to exer-cise: acute manipulations. Respir. Physiol. Neurobiol. 2006; 151: 132–140
22. J.J. Lamanca, E.M. Haymes. Effects of iron repletion on VO2mx, endurance, and blood lactate in women. Med. Sci. Sports Exerc. 1993; Vol.25, No. 12: 1386-1392
23. B. Ekblom, A.N. Goldbarg, B. Gullbring. Response to exercise after blood loss and reinfusion. Journal of Applied Physiology. 1972; 33: 175–180
24. J.A. Calbet, C. Lundby, M. Koskolou, R. Boushel. Importance of hemoglobin concentration to exer-cise: acute manipulations. Respir. Physiol. Neurobiol. 2006; 151: 132–140
25. F.J. Buick et al. Effect of induced erythocuthemia on aerobic work capacity. Journal of Applied Physiology 1980; 48: 636-642
26. D. Costill, S. Trappe. Running: The athlete within. 2002; Traverse City, MI: Cooper Publishing Group.
27. J.A. Calbet, C. Lundby, M. Koskolou, R. Boushel. Importance of hemoglobin concentration to exercise: acute manipulations. Respir Physiol Nerubiol. 2006; 151(2-3), 132–140.
28. C.M. Beall,M.J. Decker, G.M. Brittenham, I. Kushner, A. Gebremedhin, K.P. Strohl. An Ethiopian pattern of human adaptation to high-altitude hypoxia. Proc Natl Acad Sci; 2002, 99(26), 17215–17218.
29. D.R. Bassett, E.T. Howley. Limiting factors for maximum oxygen uptake and determinants of endurance performance. Medicine and Science in Sports and Exercise. 2000; 32, 70–84
30. J.M. Hagberg, W.K. Allen, D.R. Seals, B.H. Hurley, A.A. Eshani, and J.O. Holloszy. A hemodynamic comparison of young and older endurance athletes during exercise. J. Appl. Physiol. 1985; 58:2041-2046.
31. J.H. Wilmore, P.R. Stanforth, J. Gagnon, T. Rice, S. Mandel, A.S. Leon, D.C. Rao, S. Skinner, & C. Bouchard. Cardiac output and stroke volume changes with endurance training: The heritage family study. Med Sci Sports Exerc. 2001; 22(1): 99-106.
32. J. Henriksson, J.S. Reitman. Time course of changes in human skeletal muscle succinate dehydro-genase and cytochrome oxidase activities and maximal oxygen uptake with physical activity and inactivity. Acta Physiol. Scand. 1977; 99, 91–97